De música clásica, buen humor y amígdalas extendidas
Table of Contents

Se ha reconocido que bien por Bach, Beethoven o Mozart, la música clásica puede afectar nuestro estado de ánimo. En un estudio publicado en la revista Cell Reports, científicos chinos explican cómo utilizaron mediciones de ondas cerebrales y técnicas de imágenes neuronales para mostrar de qué manera la música clásica occidental provoca efectos positivos en el cerebro. El objetivo de los investigadores es encontrar formas más efectivas de utilizar la música para activar el cerebro en aquellos que no responden de otro modo, como las personas con depresión resistente al tratamiento.
“Nuestra investigación integra los campos de las Neurociencias, la Psiquiatría y la Neurocirugía, proporcionando una base para cualquier investigación centrada en la interacción entre la música y las emociones”, señaló el autor principal del paper, Bomin Sun, director y profesor del Centro de Neurocirugía Funcional de la Universidad Jiao Tong de Shanghai. “En última instancia, esperamos traducir los hallazgos de nuestra investigación a la práctica clínica, desarrollando herramientas y aplicaciones de musicoterapia que sean convenientes y efectivas”.
Depresión resistente al tratamiento #
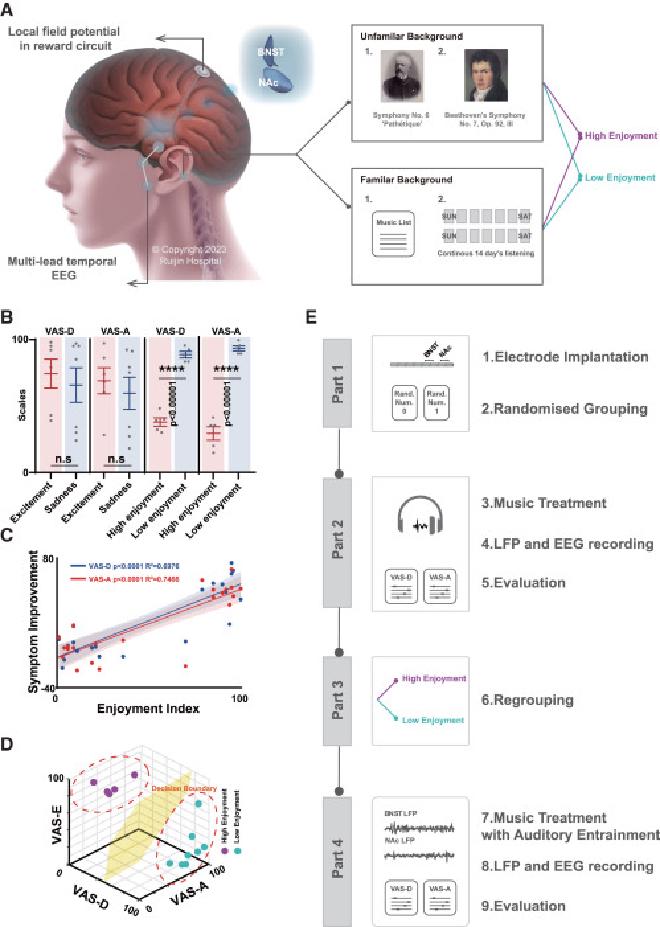
(A) Los pacientes se sometieron a un tratamiento musical mientras grababan LFP en el circuito BNST-NAc y señales temporales de EEG. Primero se dividieron aleatoriamente veintitrés sujetos en dos grupos con antecedentes de familiaridad diferenciados. Para el fondo de la UF, 13 de 23 sujetos recibieron música de la UF que representaba distintos colores emocionales. El primer grupo se llamó “tristeza” y el segundo grupo se llamó “emoción”. Los 10 sujetos restantes recibieron música de fondo familiar (FA), que se definió como reproducida continuamente durante 14 días para lograr un nivel alto y cercano de familiaridad entre ellos. Después de 14 días, los participantes se dividieron aleatoriamente en dos grupos. Un grupo escuchó la música que más le gustaba, mientras que el otro grupo escuchó la música que menos le gustaba. (B) Comparación de las puntuaciones de depresión visual y ansiedad en la VAS-D y la VAS-A entre los grupos de tristeza y excitación después de escuchar música. Los resultados no mostraron diferencias significativas en los síntomas relacionados con la depresión entre los dos grupos (prueba t no apareada, n = 6 en excitación, n = 7 en tristeza, n = 5 en alto disfrute, n = 5 en bajo disfrute). La línea central del gráfico representa el valor medio, mientras que las barras de error indican el error estándar de la media (SEM). (C) Análisis de correlación entre el grado de apreciación musical y la mejora de los síntomas depresivos (n = 13). Hubo una relación de regresión lineal significativa entre la VAS-E, VAS-D y VAS-A de los pacientes antes y después de escuchar música, lo que indica que el efecto antidepresivo de la música sobre la TRD puede estar relacionado con el propio nivel de disfrute musical de los pacientes. La línea central representa el valor medio, mientras que el área sombreada indica el SEM. (D) Los pacientes se reagruparon según los resultados de la escala. Uso de puntuaciones VAS-D, VAS-A y VAS-E, combinadas con métodos de agrupación espacial y K-vecinos más cercanos para el cálculo de límites de decisión. (E) Resumen de todo el procedimiento experimental.
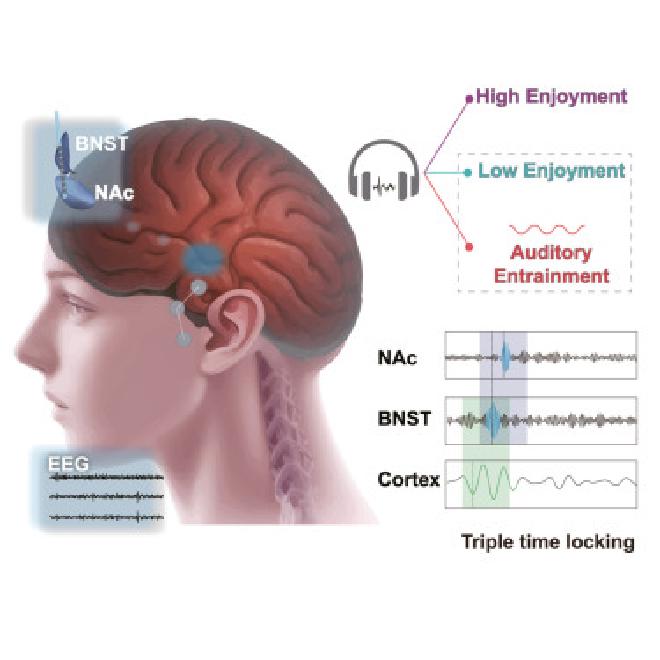
El estudio se centró en 13 pacientes con depresión resistente al tratamiento a quienes ya se les habían implantado electrodos en el cerebro con el fin de realizar una estimulación cerebral profunda. Estos implantes se colocan en un circuito que conecta dos áreas del prosencéfalo: el núcleo del lecho de la estría terminal(BNST) y el núcleo accumbens(NAc). Utilizando estos implantes, los investigadores descubrieron que la música genera efectos antidepresivos sincronizando las oscilaciones neuronales entre la corteza auditiva, que es responsable del procesamiento de la información sensorial, y el circuito de recompensas, que es responsable del procesamiento de la información emocional.
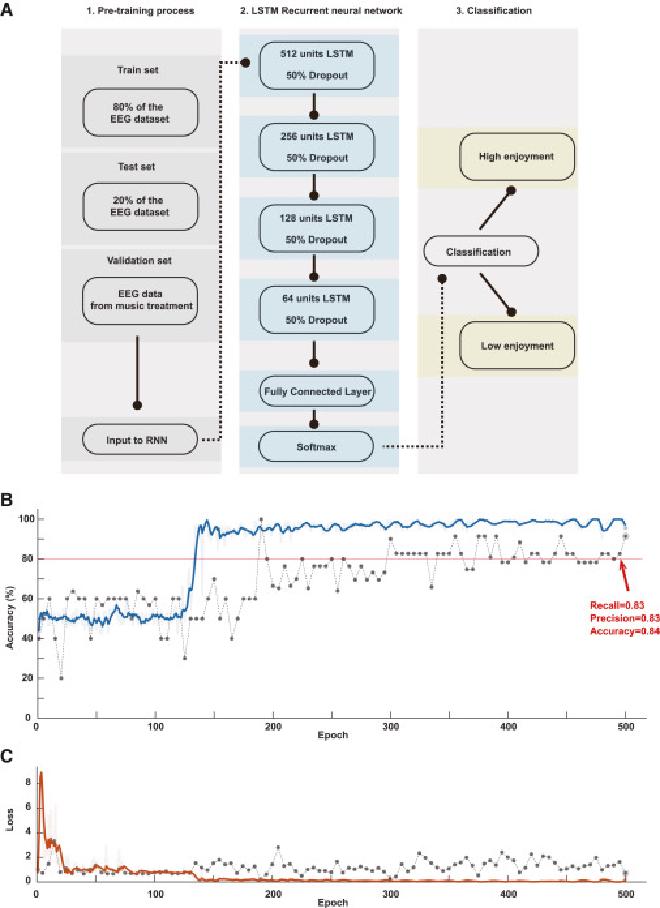
(A) Construido utilizando un algoritmo de aprendizaje profundo basado en memoria a corto plazo (LSTM), el conjunto de datos se dividió y se determinaron los resultados de salida. (B) Cambios en la precisión del conjunto de validación a medida que aumenta el número de épocas. Después de 500 épocas, se logró una precisión de 0,84, una precisión de 0,83 y una recuperación de 0,83, lo que indica que las señales temporales de EEG pueden clasificar eficazmente los subgrupos que se muestran en la Figura 1D. (C) Cambios en la pérdida en el conjunto de entrenamiento a medida que aumenta el número de épocas. Se puede observar que, a medida que el proceso de entrenamiento logra una buena precisión de clasificación, no hubo un sobreajuste significativo.
“El circuito BNST-NAc, a veces denominado parte de la ‘ amígdala extendida’1, subraya la estrecha relación entre este circuito y la amígdala, una estructura central en el procesamiento de la información emocional”, explicó Sun. “Este estudio revela que la música induce un triple bloqueo de las oscilaciones neuronales en el circuito cortical-BNST-NAc a través de la sincronización auditiva”.
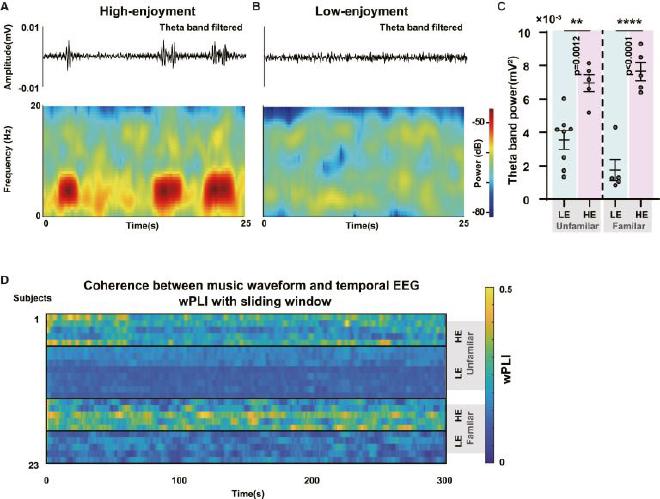
(A) Esquema de espectrogramas y formas de onda sin procesar filtradas con banda theta de EEG temporal en el sujeto del subgrupo HE. (B) Esquema de espectrogramas y formas de onda sin procesar filtradas de banda theta de EEG temporal en el sujeto LE. (C) Comparación de la potencia de la banda theta entre los subgrupos LE y HE. Los dos paneles de la izquierda representan el grupo de fondo UF, mientras que los dos paneles de la derecha representan el grupo de fondo FA. En ambos entornos de UF y FA, los subgrupos HE exhibieron un poder de banda theta significativamente mayor (n = 8 en LE UF, n = 5 en HE UF, n = 5 en LE FA, n = 5 en HE FA, ANOVA unidireccional con la prueba de Tukey). La línea central representa el valor medio, mientras que las barras de error indican el SEM. (D) Mapa de calor que ilustra la sincronización entre las formas de onda de la música y el EEG temporal.
Los pacientes del estudio fueron asignados a dos grupos: baja apreciación musical o alta apreciación musical. Aquellos en el grupo de alta apreciación musical demostraron una sincronización neuronal más significativa y mejores efectos antidepresivos, mientras que aquellos en el grupo de baja apreciación musical mostraron resultados más pobres. Al agrupar a los pacientes, los investigadores pudieron estudiar los mecanismos antidepresivos de la música con mayor precisión y proponer planes de musicoterapia personalizados que mejorarían los resultados del tratamiento. Por ejemplo, al insertar ruido de frecuencia theta en la música para mejorar el acoplamiento oscilatorio BNST-NAc, aquellos en el grupo de pacientes con baja apreciación musical informaron un mayor disfrute de la música.
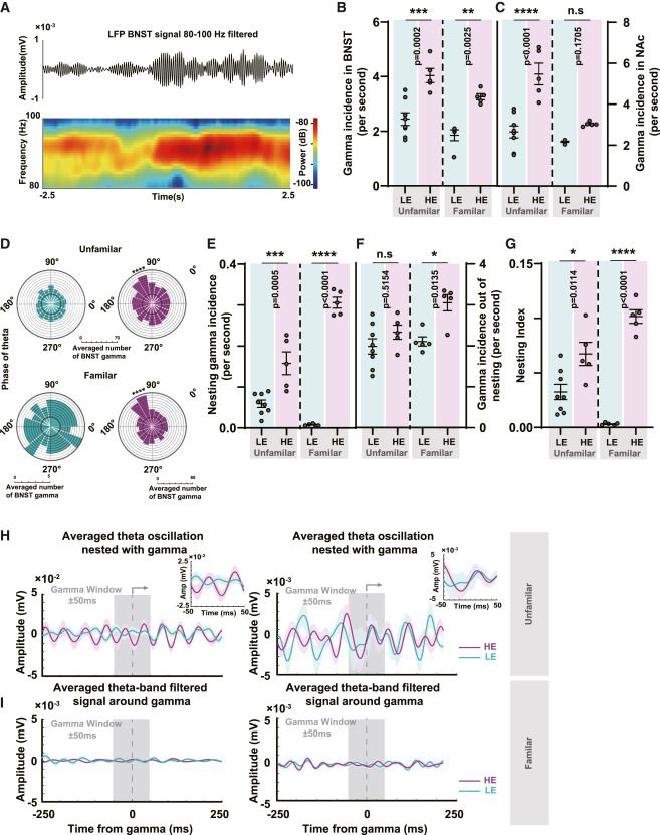
(A) Esquema de las formas de onda filtradas de la banda gamma BNST-LFP y sus espectrogramas en un sujeto del subgrupo HE. (B) Comparación de las tasas de aparición de oscilaciones gamma en BNST LFP entre los subgrupos LE y HE. En ambos entornos de FA y UF, los subgrupos HE tuvieron tasas de aparición de oscilación gamma significativamente más altas que el subgrupo LE (n = 8 en LE UF, n = 5 en HE UF, n = 5 en LE FA, n = 5 en HE FA , ANOVA unidireccional con la prueba de Tukey). La línea central representa el valor medio, mientras que las barras de error indican el SEM. (C) Comparación de las tasas de aparición de oscilaciones gamma en NAc LFP (n = 8 en LE UF, n = 5 en HE UF, n = 5 en LE FA, n = 5 en HE FA, ANOVA unidireccional con prueba de Tukey). La línea central representa el valor medio, mientras que las barras de error indican el SEM. (D) Tanto en el fondo de UF como de FA, análisis del bloqueo de fase de BNST gamma a theta temporal de EEG en el subgrupo HE y el subgrupo LE. BNST gamma exhibió una amplitud significativamente mayor alrededor de 120° de la fase theta temporal del EEG (n = 8 en LE UF, n = 5 en HE UF, n = 5 en LE FA, n = 5 en HE FA, prueba t no apareada). (E) Comparación de las tasas de aparición de oscilaciones gamma anidadas dentro del EEG theta temporal tanto en los subgrupos HE como en los subgrupos LE (n = 8 en LE UF, n = 5 en HE UF, n = 5 en LE FA, n = 5 en HE FA, ANOVA unidireccional con prueba de Tukey). La línea central representa el valor medio, mientras que las barras de error indican el SEM. (F) Comparación de las tasas de aparición de oscilaciones gamma no anidada dentro del EEG theta temporal (n = 8 en LE UF, n = 5 en HE UF, n = 5 en LE FA, n = 5 en HE FA, ANOVA unidireccional con la prueba de Tukey). La línea central representa el valor medio, mientras que las barras de error indican el SEM. (G) Comparación del índice de anidación, que ilustra la relación temporal de la anidación theta-gamma. En ambos entornos de UF y FA, los subgrupos HE mostraron tasas de aparición de oscilaciones gamma significativamente más altas que los subgrupos LE (n = 8 en LE UF, n = 5 en HE UF, n = 5 en LE FA, n = 5 en HE FA , ANOVA unidireccional con la prueba de Tukey). La línea central representa el valor medio, mientras que las barras de error indican el SEM. (H) En ambos entornos de UF y FA, las formas de onda theta del EEG alrededor de la amplitud del pico gamma anidado promediaron 250 ms antes y después, revelando una modulación oscilatoria más rítmica en el subgrupo HE (para la izquierda, n = 237 en UF HE, n = 144 en UF LE; para la derecha, n = 522 en UF HE, n = 3 en UF LE). La región gris semitransparente, que abarca 50ms antes y después del tiempo máximo de gamma, representa la ventana de actividad de oscilación gamma. Las líneas representan la forma de onda theta media y el área sombreada indica la desviación estándar. (I) En el grupo de fondo de UF, la amplitud promedio del pico gamma no anidado antes y después de 250 ms de datos filtrados con banda theta de EEG no mostró diferencias significativas entre los dos grupos (n = 3492 en el subgrupo HE, n = 4088 en el subgrupo LE). De manera similar, en el grupo de antecedentes FA, el promedio tampoco mostró diferencias significativas entre los dos subgrupos (n = 5193 en el subgrupo HE, n = 58 en el subgrupo LE).
Por qué música clásica #
_En el estudio se utilizaron varias piezas de música clásica occidental. Se eligió este tipo de música porque la mayoría de los participantes no estaban familiarizados con ella y los investigadores querían evitar cualquier interferencia que pudiera surgir de la familiaridad subjetiva. “Concluimos que las elecciones musicales durante el proceso de escucha formal fueron individualizadas y no estaban relacionadas con el trasfondo emocional de la música", dijo Sun.
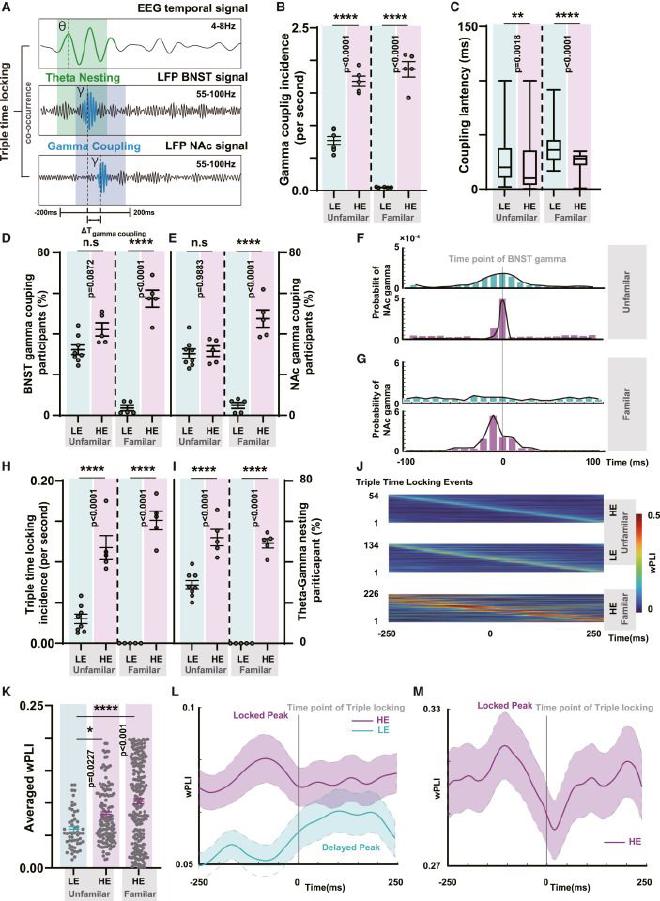
(A) Esquema del bloqueo de tiempo triple. (B) Comparación de la incidencia del acoplamiento gamma-gamma en el circuito BNST-NAc (n = 8 en LE UF, n = 5 en HE UF, n = 5 en LE FA, n = 5 en HE FA, unidireccional ANOVA con la prueba de Tukey). La línea central representa el valor medio, mientras que las barras de error indican el SEM. (C) Comparación de la latencia del acoplamiento gamma-gamma en el circuito BNST-NAc (n = 8 en LE UF, n = 5 en HE UF, n = 5 en LE FA, n = 5 en HE FA, unidireccional ANOVA con prueba de Tukey, n = 1257 para el subgrupo HE versus n = 2601 para el subgrupo LE, n = 3128 para el subgrupo HE versus n = 52 para el subgrupo LE). La línea central representa el valor medio, mientras que las barras de error indican el SEM. (D) Comparación de los participantes en el acoplamiento de oscilaciones gamma BNST (n = 8 en LE UF, n = 5 en HE UF, n = 5 en LE FA, n = 5 en HE FA, ANOVA unidireccional con prueba de Tukey). La línea central representa el valor medio, mientras que las barras de error indican el SEM. (E) Comparación de los participantes de las oscilaciones gamma NAc en acoplamiento (n = 8 en LE UF, n = 5 en HE UF, n = 5 en LE FA, n = 5 en HE FA, ANOVA unidireccional con la prueba de Tukey). La línea central representa el valor medio, mientras que las barras de error indican el SEM. (F) Relación temporal dentro de los eventos de acoplamiento gamma-gamma en el grupo de fondo UF. Utilizando la aparición de BNST-gamma como punto cero, se representó gráficamente la incidencia de NAc-gamma. Se pudo observar que el subgrupo HE exhibe una tendencia más fuerte de aparición de NAc-gamma alrededor de BNST-gamma, lo que indica que BNST-gamma recluta NAc-gamma. (G) Relación temporal dentro de los eventos de acoplamiento gamma-gamma en el grupo de fondo FA. (H) Comparación de la tasa de aparición de eventos de triple bloqueo temporal (n = 8 en LE UF, n = 5 en HE UF, n = 5 en LE FA, n = 5 en HE FA, ANOVA unidireccional con la prueba de Tukey ). La línea central representa el valor medio, mientras que las barras de error indican el SEM. (I) Comparación de los participantes de anidación theta-gamma en eventos de bloqueo de tiempo triple (n = 8 en LE UF, n = 5 en HE UF, n = 5 en LE FA, n = 5 en HE FA, unidireccional ANOVA con la prueba de Tukey). La línea central representa el valor medio, mientras que las barras de error indican el SEM. (J) Mapa de calor que muestra los resultados del análisis de sincronización entre la música y la actividad cortical auditiva dentro de la ventana de tiempo de bloqueo de tiempo peri-triple. (K) Comparación del wPLI promedio de bloqueo de tiempo peri-triple en el fondo UF de HE y LE y el fondo FA de HE (n = 54 en LE UF, n = 134 en HE UF, n = 226 en HE FA, ANOVA unidireccional con la prueba de Tukey). El promedio se realizó durante una ventana de tiempo antes y después de cada evento de triple bloqueo de tiempo, resaltando la comparación de sincronización general y mitigando las variaciones dentro de la ventana de tiempo. (L) En el grupo de fondo UF, se promedió wPLI dentro de la ventana de bloqueo de tiempo peri-triple, lo que muestra sincronización entre la música y las respuestas corticales auditivas. Hubo un pico alrededor de 50ms antes del bloqueo de tiempo triple (n = 134 en UF HE y n = 54 en UF LE). La línea central representa el valor medio, mientras que el área sombreada indica el SEM. (M) En el grupo de fondo UF, se promedió wPLI dentro de la ventana de bloqueo de tiempo peri-triple (n = 226 en FA HE). La línea central representa el valor medio, mientras que las barras de error indican el SEM.
La investigación futura del equipo se centrará en varias áreas. Por un lado, su objetivo es estudiar cómo la interacción entre la música y las estructuras profundas del cerebro desempeña un papel en los trastornos depresivos. También introducirán otras formas de estímulos sensoriales, incluidas imágenes visuales, para investigar los posibles efectos terapéuticos combinados de la estimulación multisensorial sobre la depresión.
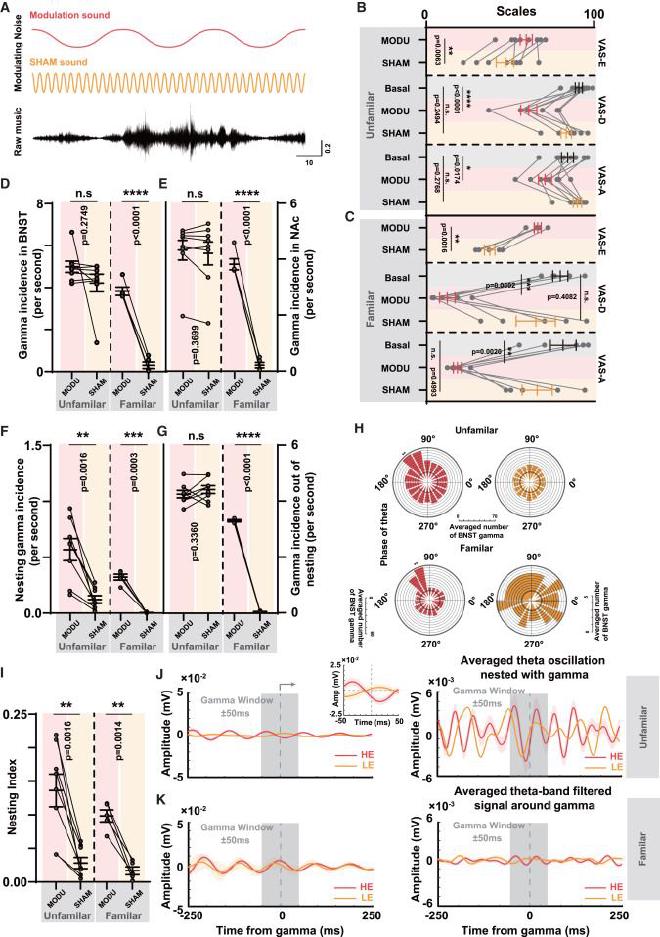
(A) Esquema del sonido modulador superpuesto a la forma de onda original. La línea roja representa la forma de onda del arrastre auditivo, mientras que la línea amarilla representa la forma de onda del ruido SHAM. La línea negra representa la forma de onda original de la música. (B) Comparación de los resultados de tres escalas de calificación visual en el fondo de la UF. En la parte superior se muestra la comparación de las calificaciones VAS-E cuando los pacientes escucharon ambos tipos de música (n = 8, prueba t pareada), en el centro están las calificaciones VAS-D (n = 8, análisis de varianza de medidas repetidas [ RM-ANOVA] con la prueba de Dunnett), y en la parte inferior están las calificaciones VAS-A (RM-ANOVA con la prueba de Dunnett). (C) Comparación de los resultados de tres escalas de calificación visual en el fondo de FA (n = 5, prueba t pareada y RM-ANOVA con prueba de Dunnett). La línea central del gráfico representa el valor medio, mientras que las barras de error indican el SEM. (D) Comparación de las tasas de aparición de BNST-gamma (n = 8, prueba t pareada). La línea central del gráfico representa el valor medio, mientras que las barras de error indican el SEM. (E) Comparación de las tasas de aparición de NAc-gamma (n = 5, prueba t pareada). La línea central del gráfico representa el valor medio, mientras que las barras de error indican el SEM. (F) Comparación de la incidencia de anidación de theta-gamma (n = 8, prueba t pareada). La línea central del gráfico representa el valor medio, mientras que las barras de error indican el SEM. (G) Comparación de la incidencia de aparición de gamma no anidada (n = 5, prueba t pareada). La línea central del gráfico representa el valor medio, mientras que las barras de error indican el SEM. (H) En ambos fondos de UF y FA, análisis de bloqueo de fase de BNST gamma a theta temporal de EEG en el subgrupo MODU y el subgrupo SHAM. BNST gamma exhibió una amplitud significativamente mayor alrededor de 120° de la fase theta temporal del EEG (n = 8 en UF y n = 5 en FA, prueba t no apareada). (I) Comparación del índice de anidación (n = 8 en UF y n = 5 en FA, prueba t no apareada). (J) En el grupo de fondo UF (izquierda), las formas de onda theta promedio de EEG alrededor de la amplitud del pico gamma anidado muestran una modulación más rítmica en el subgrupo MODU (n = 1093) y el subgrupo SHAM (n = 236). En el grupo de fondo FA, el subgrupo MODU (n = 522) fue comparable al grupo UF, mientras que el grupo SHAM solo encontró un evento de bloqueo de tiempo triple (n = 1). La región gris semitransparente, que abarca 50ms antes y después del tiempo máximo de gamma, representa la ventana de tiempo de la actividad de oscilación gamma. (K) En el grupo de fondo UF, no hubo diferencias significativas entre el subgrupo MODU (n = 8159) y el grupo SHAM (n = 8471) en los datos filtrados promedio de la banda theta de EEG alrededor de la amplitud del pico gamma no anidado. En el grupo de antecedentes de FA, se hizo la misma comparación con el subgrupo MODU (n = 5193) y el grupo SHAM (n = 72). Las líneas del gráfico representan la forma de onda theta media y el área sombreada representa la desviación estándar.
Teléfonos inteligentes #
“Al colaborar con médicos, músicoterapeutas, informáticos e ingenieros, planeamos desarrollar una serie de productos de salud digitales basados en musicoterapia, como aplicaciones para teléfonos inteligentes y dispositivos portátiles", señaló Sun. “Estos productos integrarán recomendaciones musicales personalizadas, monitoreo y retroalimentación emocional en tiempo real y experiencias multisensoriales de realidad virtual para brindar herramientas de autoayuda convenientes y efectivas para controlar las emociones y mejorar los síntomas en la vida diaria”.
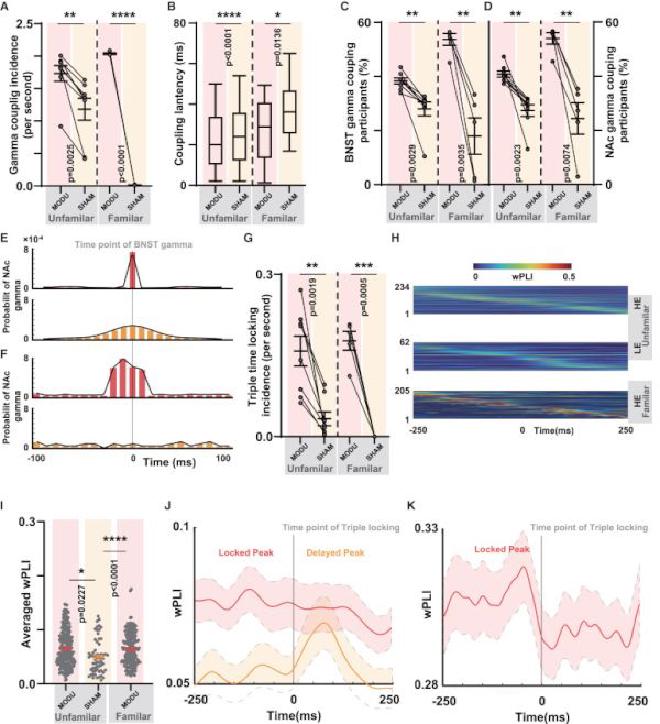
(A) Comparación de la incidencia del acoplamiento gamma-gamma en el circuito BNST-NAc tanto en los subgrupos MODU como en los subgrupos SHAM. En ambos entornos, los subgrupos MODU muestran una incidencia de acoplamiento gamma significativamente mayor (prueba t pareada, n = 8, p = 0,0025 para MODU UF versus SHAM UF; prueba t pareada, n = 5, p = 0,0001 para MODU FA versus SHAM FA). (B) Comparación de la latencia del acoplamiento gamma-gamma en el circuito BNST-NAc (ANOVA unidireccional, n = 1904 en MODU UF versus n = 1416 en SHAM UF, p0,0001; n = 2962 en MODU FA versus n = 17 en SHAM FA, p = 0,0136). (C) Comparación de los participantes de oscilación gamma BNST en eventos de acoplamiento gamma (prueba t pareada, n = 8, p = 0,0029 para MODU UF versus SHAM UF; prueba t pareada, n = 5, p = 0,0035 para MODU FA versus SHAM FA). (D) Comparación de los participantes de la oscilación gamma NAc en eventos de acoplamiento gamma (prueba t pareada, n = 8, p = 0,0023 para MODU UF versus SHAM UF; prueba t pareada, n = 5, p = 0,0074 para MODU FA versus SHAM FA). (E) Relaciones temporales internas dentro de los eventos de acoplamiento gamma tanto en el subgrupo MODU como en el subgrupo SHAM en el contexto de UF. (F) Relaciones temporales internas dentro de los eventos de acoplamiento gamma tanto en el subgrupo MODU como en el subgrupo SHAM en el contexto de FA. (G) Comparación de la tasa de aparición de eventos de bloqueo de tiempo triple (prueba t pareada, n = 8, p= 0,0019 para MODU UF versus SHAM UF; prueba t pareada, n = 5, p = 0,0005 para MODU FA versus SHAM FA ). (H) Mapa de calor que muestra los resultados del análisis de sincronización entre la música y la actividad cortical auditiva dentro de la ventana de tiempo de bloqueo de tiempo peri-triple. Los tres paneles de arriba a abajo representan el fondo UF de MODU y SHAM y el fondo FA de MODU. (I) Comparación del wPLI promedio de bloqueo de tiempo peri-triple en el fondo UF de MODU (n = 134) y SHAM (n = 54), y el fondo FA de MODU (n = 205) (ANOVA unidireccional con Prueba de Tukey, UF MODU versus UF SHAM, p = 0,0227, UF SHAM versus FA MODU, p 0,0001). (J) En el grupo de fondo de UF, el wPLI se promedia en cada punto de tiempo dentro de la ventana de tiempo de bloqueo de tiempo peri-triple, destacando la tendencia de sincronización entre la música y las respuestas corticales auditivas dentro de la ventana de tiempo (n = 134 en UF MODU y n = 54 en UF SHAM). En el grupo de fondo de FA, el wPLI se promedia en cada punto de tiempo dentro de la ventana de tiempo de bloqueo de tiempo peri-triple (n = 205 en FA MODU). Las líneas representan la forma de onda theta media y el área sombreada representa la desviación estándar.
Importante #
-
El estudio Auditory entrainment coordinates cortical-BNST-NAc triple time locking to alleviate the depressive disorder fue publicado en Cell Reports. Sus autores son: Xin Lv, Yuhan Wang, Yingying Zhang, Shuo Ma, Jie Liu, Kuanghao Ye, Yunhao Wu, Valerie Voon & Bomin Sun.
-
El estudio contó con el apoyo de la National Natural Science Foundation of China, la Shanghai Jiao Tong University, el plan de acción de innovación científica y tecnológica de Shanghai, y el Shanghai Municipal Science and Technology Major Project
-
Las imágenes en este artículo, acreditadas a Bomin et al., forman parte del paper Auditory entrainment coordinates cortical-BNST-NAc triple time locking to alleviate the depressive disorder, publicado en Cell Reports
Contacto - Contact Us #
A menudo tengo la inquietud de saber si los autores de los estudios, papers, revisiones, investigaciones que compartimos, traducimos aquí, tendrán alguna sugerencia, corrección, contribución para hacer. Tal vez, hay una actualización de esos estudios y papers, y quieren compartirla con nosotros y que la publiquemos. Si se preguntó si leeremos esos artículos -temáticas afines a las abordamos en Notaspampeanas- por favor, utilicen el correo electrónico que está un poco más abajo. ¡Gracias!
I often have the concern of knowing if the authors of the studies, papers, reviews, researchs that we share, translate here, will have any suggestions, corrections, or contributions to make. Perhaps, there is an update to those studies and papers, and they want to share it with us and have us publish it. If you were wondering if we will read those articles - topics similar to those we address in Notaspampeanas - please use the email address below. Thank you!
Do You Like This? Make a donation - ¿Te gusta esto? Doná #
Donaciones-Donations
Alias CBU: ALETA.GRIS.CORDON
BTC: 3GqfXZorhmypEAPrhf8LuVhfgbJWziWk7N
ETH: 0x9F036AC4FCc1F025e1B5e9dA438826E310B20245
USD Coin: 0x9F036AC4FCc1F025e1B5e9dA438826E310B20245
Ltc: MJqeqEsx6QDPpfDCXdNJEE2VEBUmWk9nQr
USDT: 0x9F036AC4FCc1F025e1B5e9dA438826E310B20245
DOGE: DMh6sDEFbY9QVsK6x3f2952wWzG2ty89Rh
-
La amígdala extendida es una estructura que se encuentra en el cerebro de los mamíferos, incluyendo a los seres humanos. Esta región es conocida por su papel en la regulación de las emociones y el procesamiento del miedo, así como en la motivación y la toma de decisiones. Además de su implicación en las respuestas emocionales, también juega un papel importante en la adicción y el estrés crónico. La amígdala extendida está compuesta por varias regiones interconectadas que desempeñan distintas funciones relacionadas con el procesamiento emocional y motivacional. ↩︎